Singing Humpback Whales Associate with Mothers and Calves
Marsha L. Green 1, 2 Ann M. Zoidis 1 Jeff Pantukhoff 3
1) Ocean Mammal Institute, Po Box 14422, Reading, PA 19612
2) Albright College, PO Box 15234, Reading, PA 19612
3) Whaleman Foundation, PO Box 1670 Lahaina, HI 96767
Abstract
Singing humpback whales (Megaptera novaeangliae) have been well documented in Hawaiian waters between the months of January and April. Previous research indicates that singers usually are alone (Winn and Winn, 1978; Tyack, 1981; Darling, 1983). However, the occurrence of singers with other whales has been documented (Baker and Herman 1984; Helweg et al., 1990). The acoustic repertoire of humpback whales was studied off Maui, Hawaii in 1996, 1997 and 2001 by opportunistically approaching pods in a small vessel. Both singing (rhythmic and continuous sounds) and social sounds (non-rhythmic, without a consistent pattern) were documented. Over the 3 years of data collection 43 of the 330 pods approached had singers. For twenty-five of the singing pods, vocalizations were recorded using underwater video photography with a self-contained hydrophone in order to assess pod composition and behavior. For eighteen of the singing pods, recordings were done with an underwater hydrophone suspended from the research vessel. Of the 25 pods observed underwater, singers were with a mother and calf 68% of the time (n=17). The remaining 8 singers observed underwater were alone.
These results indicate that singing humpback whales may be associated with mothers and calves more frequently than previously documented. While the function of humpback whale song remains uncertain (see Helweg et al. for a discussion of possible purposes of song), these results suggest there may be some functional significance to singers associating with females with calves.
Introduction
"Humpback whale song has been described as the most elaborate single display of any animal species" (Wilson, 1975). The song was initially described in detail by Payne and McVay (1971) and Winn, Perkins and Poulter (1971). Singing whales most often are reported to be alone (Winn and Winn, 1978; Tyack, 1981) although there are exceptions (Baker and Herman, 1984; Frankel, et al., 1989). It is generally accepted that singing is done by males where calves are being born and seasonal gonadal activity is high (Chittleborough, 1955; Payne and McVay, 1971). The relationship between singing and seasonal gonadal activity suggests that song production plays a role in the mating system. While humpback whale song appears to be the result of strong sexual selection (Tyack, 1981), its function in the mating system remains unclear.
Methods
In 1996, 1997 and 2001 we opportunistically approached humpback whale pods in a small vessel in the area between Maui, Lanai and Kahoolawe, Hawaii. Both singing and social sounds (non-rhythmic, without a consistent pattern) were documented. When sea state and water clarity were favorable, vocalizations were recorded using underwater video photography with a self-contained hydrophone in order to assess pod composition and behavior. In other instances vocalizations were recorded with a hydrophone suspended from the research vessel.
Results
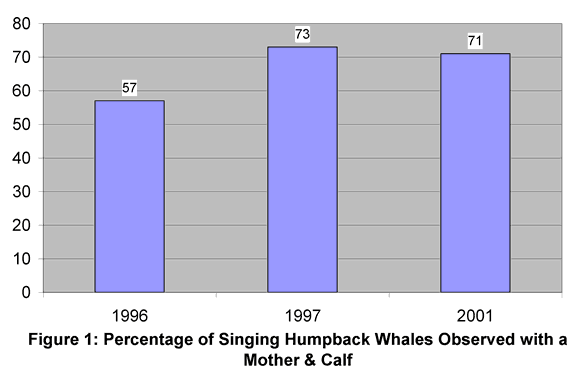
Over the three years we approached 330 pods. Forty-three of these pods had singers. A researcher entered the water to videotape twenty-five of the pods that had a singing whale. Seventeen of these twenty-five singers were associated with a mother and calf. The other eight pods observed underwater had lone singers. See Figure 1 for percentage of singers associated with a mother and calf over three years of observation. Figure 2 gives the percentage of mother/calf escort pods where the escort was singing.
Discussion
Other researchers have observed singing whales with mothers and calves (Tyack and Whitehead, 1983; Baker and Herman, 1984; Helweg et al., 1990). However, it is generally accepted that singers tend to be alone (Winn and Winn, 1978; Tyack, 1981). The present study found that the majority of the singers observed underwater were escorts, indicating that singing humpback whales may be associated with mothers and calves more frequently than previously documented. Since this data was collected in January, February, March and April over three years, it is not likely that the results were skewed by unusual circumstances in any one year.
Since we opportunistically approached whales we don’t know if the mother and calf joined the singer or vice versa. Our underwater observations indicate the mother and calf were not attempting to swim away from the singer, but were allowing the presence of the singing escort. They were either swimming slowly or remaining relatively stationary (possibly resting) with the exception of one pod that was swimming moderately fast. Tyack (1981) reported that when a singer joined with a mother and calf, behavior tentatively associated with courtship and mating was observed. We did not observe any tentative courtship or mating behavior.
Various possibilities have been discussed about the function of male humpback whales escorting females with calves. Mobley and Herman (1985) suggest that males may associate with mothers with newborn calves to see if the female is ovulating postpartum and that, if copulation occurs, the male may engage in postcopulatory mate guarding.
Various theories also have been proposed about the function of humpback whale song. One theory is that singing may be a male sexual display to attract females (Payne and McVay, 1971, Winn and Winn, 1998; Tyack, 1981). In this case females select which male to associate with based on the characteristics of his song (loudness, duration, etc). Others suggest that song may establish dominance among males or establish or maintain space between competing males (Winn and Winn, 1978; Tyack, 1981; Frankel and Herman, 1987; Mobley et al., 1988). Humpback whale song could, perhaps, function in both female choice and male/male competition.
The observation that 68% of singing humpback whales observed underwater were with a mother and calf appears to support the theory that the female may use the song to select which male to associate with, i.e., the song functions to attract females for reproductive purposes. However, it appears that most of the actual reproduction may not take place immediately (while the mother is with her calf) since most females do not have calves every year.
Research indicates that female humpback whales produce one calf an average of every 2.4 to 2.8 years (Chittleborough, 1958, 1965; Baker et al., 1987; Clapham and Mayo, 1987). However, some mothers with newborn calves may ovulate postpartum (Chittleborough, 1958). If song functions mainly to attract females for mating purposes, why would such a high percentage of singing whales spend time with females with calves, since the probability of a female with a calf mating and becoming pregnant is relatively low? Perhaps the mother and escort are developing social bonds or reinforcing existing bonds for possible reproduction in subsequent years.
Some male pied kingfishers without mates are known to help an unrelated nesting pair raise their young because it ultimately increases their direct fitness by increasing the probability that the female they assisted will mate with them the next year (Reyer, 1984). Possibly the female humpback whale uses the song to help select the optimal escort to assist her in protecting her calf from sharks and other predators or keeping other male humpback whales away. The song also may function as a signal to other males to stay away from the mother and calf. Tyack (1981) noted that whales usually turn away from singers.
Whether the song helps the female select the optimal escort for protection, or the song functions to keep other males away, or both, the protective assistance provided by the escort could form the basis of a social bond with the mother that increases the probability of the escort reproducing with that female in subsequent years. Thus, singing may function to attract females without calves for immediate mating, while singing and escorting a female with a calf may primarily enhance the males’ direct fitness in the future.
Conclusions
Acknowledgements: This data was collected under permits # 883 and #924-1484-01
from the National Marine Fisheries Service.
References
Baker, C . S., Perry, A., and Herman, L. M. 1987. Reproductive histories of female humpback whales Megaptera novaeangliae in the North Pacific. Marine Ecology Progress Series, 41:103-114.
Baker, C. S. and Herman, L. M. 1989. Behavioral responses of summering humpback whales to vessel traffic: experimental and opportunistic observations. Final Report to the National Park Service, U. S. Department of the Interior, Anchorage, AK.
Chittleborough, R. G. 1955. Aspects of reproduction in the male humpback whale, Megaptera nodosa (Bonna-terre). Australian Journal of Marine and Freshwater Research, 6:1-29.
Chittleborough, R. G. 1958. The breeding cycle of the female humpback whale Megaptera nodosa (Bonna-terre). Australian Journal of Marine and Freshwater Research, 9:1-18.
Chittleborough, R. G. 1965. Dynamics for 2 populations of the humpback whale, Megaptera novaeangliae (Borowski). Australian Journal of Marine and Freshwater Research, 16:33-128.
Clapham, P. and Mayo, C. A. 1987. Reproduction and recruitment of individually identified humpback whales, Megaptera novaeangliae, observed in Massachusetts Bay, 1979-1985. Canadian Journal of Zoology, 65:2853-2863.
Darling, J. D. 1983. Migrations, abundance and behavior of Hawaiian humpback whales,
Megaptera novaeangliae (Borowski). PhD dissertation, University of California Santa Cruz, Santa Cruz, CA.
Darling, J. D., and Jurasz, C. M. 1983. Migratory destinations of North Pacific humpback whales (Megaptera novaeangliae). in: "Communication and behavior of whales," R. Payne ed., pp. 359-368. Westview press: Boulder, CO.
Frankel, A. S., Clark, C. W., Herman, L. M., Gabriele, C. M., Hoffhines, M. A., Freeman, T. R., and Patterson, B. K. 1989. Acoustic location and tracking of wintering humpback whales (Megaptera novaeangliae) off South Kohala, Hawaii. in: "Proceedings of the Eighth Biennial Conference on the Biology of Marine Mammals," Monterey, CA. Abstracts.
Frankel, A. S. and Herman, L. M. 1987. Behavioral responses of humpback whales (Megaptera novaeangliae) to playback of sound. in: ""Proceedings of the Seventh Biennial Conference on the Biology of Marine Mammals," Miami, FL. Abstracts.
Helweg, D.A., Herman, L.M., Yamamoto, S. and Forestell, P.H. 1990. Comparison of songs of humpback whales (Megaptera novaeangliae) recorded in Japan, Hawaii, and Mexico during the winter of 1989. Scientific Reports to the Cetacean Research Institute, 1:1-12.
Mobley, J. R. and Herman, L. M. 1985. Transience of social affiliations among humpback whales (Megaptera novaeangliae) on the Hawaiian wintering grounds. Canadian Journal of Zoology, 63:762-772.
Mobley, J. R., Herman, L. M. and Frankel, A. S. 1988. Responses of wintering humpback whales (Megaptera novaeangliae) to playback of recordings of winter and summer vocalizations and of synthetic sound. Behavioral Ecology and Sociobiology, 23:211-223.
Payne, R.S and McVay, S. 1971. Songs of the humpback whales. Science. 173:585-597.
Reyer, H. U. 1984. Investment and relatedness: A cost/benefit analysis of breeding and helping in the pied kingfisher. Animal Behavior 32:1163-1178.
Tyack, P. 1981. Interactions between singing Hawaiian humpback whales and conspecifics nearby. Behavioral Ecology and Sociobiology. 8(2):105-116.
Tyack, P. and Whitehead, H. 1983. Male competition in large groups of wintering humpback whales. Behavior, 83:132-154.
Wilson, E. O. 1975. "Sociobiology." Belknap: Cambridge.
Winn, H. E., Perkins, P. J. and Poulter, T. C. 1971. Sounds of the humpback whale. in: "Proceedings of the Seventh Annual Conference on Biological SONAR and diving Mammals," pp. 39-52. Stanford Research Institute: Menlo Park, CA.
Winn, H.E. and L.K. Winn. 1978. The song of the humpback whale (Megaptera novaeangliae) in the West Indies. Marine Biology. 47:97-114.